Volume 12, Number 2—February 2006
Dispatch
Novel Dengue Virus Type 1 from Travelers to Yap State, Micronesia
Abstract
Dengue virus type 1 (DENV-1), which was responsible for the dengue fever outbreak in Yap State, Micronesia, in 2004, was isolated from serum samples of 4 dengue patients in Japan. Genome sequencing demonstrated that this virus belonged to genotype IV and had a 29-nucleotide deletion in the 3´ noncoding region.
Dengue virus (DENV) is a mosquitoborne flaviviruses; there are 4 serotypes, DENV-1, -2, -3 and -4. DENV has been found in >100 countries and 2.5 billion people live in areas where dengue is endemic. Fifty to one hundred million cases of dengue infection are estimated to occur annually (1). In Japan, outbreaks of dengue fever occurred in Nagasaki, Hiroshima, Kobe, and Osaka from 1942 to 1945, but none thereafter (2). However, ≈50 imported dengue cases occur annually in Japan.
The DENV genome is a single-stranded positive-sense RNA of ≈11,000 nucleotides (nt) that encodes 3 structural proteins (capsid, membrane, and envelope) and 7 nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) (3). Surrounding the open reading frame (ORF) are 5´ and 3´ noncoding regions (NCRs) that form RNA secondary structures (4). These regions are ≈100 and ≈400 nt, respectively (5).
Dengue fever developed in 9 Japanese patients in 2004 after they returned from Yap state. We report the genetic characterization of RNA from DENV-1 isolates from these patients.
Yap is the westernmost state of the Federated States of Micronesia and composed of 4 major islands. Yap has a total area of 102 km2 and a population of 11,241 (2000 census). The climate is moderate and fairly constant. The mean annual temperature is 27°C. Relative humidity ranges from 65% to 100% (annual mean 83%). Rainfall averages 120 inches a year and is seasonal.
In 1995, a dengue epidemic caused by DENV-4 occurred in Yap state (6), but no dengue outbreaks have since been reported. However, the Yap Epinet Team reported a dengue outbreak caused by DENV-1 in Yap state that began in the last week of May 2004. A total of 658 reported dengue fever cases (defined by the World Health Organization) occurred as of December 29, 2004. No deaths or dengue hemorrhagic fever/dengue shock syndrome cases were reported (7).
Fever, headache, and diarrhea developed in 7 Japanese adults who visited Yap after their return to Japan in August 2004. DENV infection was serologically confirmed in 5 patients (patients 1–5) by an immunoglobulin M (IgM) capture enzyme-linked immunosorbent assay (ELISA) (Focus Diagnostics Inc., Herndon, VA, USA) and an IgG ELISA (PANBIO Ltd., Brisbane, Queensland, Australia) at the National Institute of Infectious Diseases in Tokyo, Japan. Of these 5 patients, 4 had a primary DENV infection and 1 had a primary dengue infection and a secondary flavivirus infection. DENV infection was serologically confirmed in the sixth patient at another institute. The seventh patient did not visit a medical facility but had symptoms of dengue fever. In addition, 2 other Japanese patients who traveled to Yap in September 2004 were diagnosed with dengue (data not shown).
Four virus isolates (NIID04-27, -31, -41, and -47) were obtained from serum samples from patients 1–4, respectively. Two hundred microliters of serum samples diluted 1:40 was injected onto C6/36 cells in minimal essential medium supplemented with 2% fetal calf serum. The cells were incubated at 28°C for 7 days and culture supernatant fluids were collected. DENV isolates were used for analysis without any further passage.
Complete nucleotide sequencing of RNA of NIID04-27, -31, and -47 and partial sequencing of NIID04-41 were performed. Viral RNA was extracted by using a High Pure RNA extraction kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions, transcribed to cDNA, and amplified by polymerase chain reaction, as described previously (8). The cDNA was purified and sequenced by using the ABI PRISM 3100 Avant Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). Seventeen pairs of primers were designed based on the DENV-1 NIID02-20 sequence (GenBank accession no. AB178040) and used in the analyses (9).
The nucleotide sequences of the viral isolates were compared with published complete sequences of DENV-1 (Table 1). Sequence alignment and analysis were performed by using ATGC analysis programs (version 4.02; Genetyx Corp., Tokyo, Japan). Phylogenetic analyses of nucleotide sequences were conducted with ClustalX software version 1.83 (ftp://ftp-igbmc.u-strasbg.fr/pub/ClusterW/). A phylogenetic tree was reconstructed for aligned nucleotide sequences by using the neighbor-joining method. Bootstrap reassembling analysis of 1,000 replicates was used to assess confidence values for virus groupings. The phylogenetic tree was constructed by using Treeview software version 1.6.6 (http://taxonomy.zoology.gla.ac.uk/rod/treeview.htm).
The full-length RNA genomes of NIID04-27, -31, and -47 were 10,706 nt. A previous study reported that the full-length RNA genome of DENV-1 was 10,735 nt (8). The differences in the genome sequence between NIID04-27 and the other 2 isolates (NIID04-31 and -47) were subtle; identities with NIID04-31 and -47 were 99.94% and 99.92%, respectively. The results suggest that these 3 isolates belong to the same strain. Therefore, we used NIID04-27 as a representative isolate for further analysis.
Figure 1
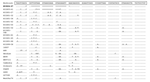
Figure 1. Nucleotide sequence alignment of the variable region in the 3´ noncoding region of dengue virus type 1 strains, including NIID04-27 sequenced in the present study. The Mochizuki strain was used as...
To characterize the molecular structure of the genome, the complete NIID04-27 nucleotide sequence was compared with those of other DENV-1 strains available in GenBank (Table 1). NIID04-27 shared sequence identity ranging from 90.9% to 96.9% (Table 2) with 12 other DENV-1 strains. With respect to the alignment of full-length genomes, some alterations were found in the 3´ NCR. These alterations included a deletion of 29 nt starting at the 13th position from the ORF termination codon (Figure 1). The same deletion in the 3´ NCR was found in the viral genome amplified directly from the serum sample from patient 1 and was also observed in NIID04-31, -41, and -47.
To further analyze the genetic variation in the 3´ NCR of DENV-1, we analyzed the sequence of 24 other DENV-1 strains. Only the NIID03-41 strain, which was isolated in our laboratory from a patient returning from the Republic of Seychelles, had a 17-nt deletion in the 3´ NCR (Figure 1). The complete genomes of the 25 DENV-1 strains analyzed showed high levels of nucleotide sequence identity in the 3´ NCR, except for a small region of 50 nt immediately after the ORF, which is the hypervariable region. The nucleotide sequence identities in the 3´ NCR between NIID04-27 and 12 other DENV-1 strains ranged from 89.3% to 92.5% (Table 2).
Figure 2

Figure 2. Phylogenetic tree based on the full-length genome sequence of 21 available dengue virus (DENV) type 1 strains and DENV-2, -3, and -4. The multiple sequence alignments were obtained with ClustalX, and...
To understand the genetic relationships and evolution of DENV-1 strains, we also performed phylogenetic analysis of the fully sequenced DENV-1 strains that included NIID04-27 (Figure 2). NIID 04-27 belonged to genotype IV along with A88, 98901518, 98901530, NIID03-41 and West Pac74 .This cluster was called the Pacific group in a previous report (10). NIID04-27 and NIID03-41 are the first DENV-1 strains to have deletions in 3´ NCR.
We have genetically characterized DENV-1 isolate NIID04-27 by determining its complete nucleotide sequence and comparing the sequence with most of the available DENV-1 full-length sequences. Sequence heterogeneity in the 3´ NCR of the genus Flavivirus has been reported for tickborne encephalitis virus, Japanese encephalitis virus, DENV-2, and DENV-4 (11–13). For example, DENV-2 isolated in Texas, Peru, Venezuela, Mexico, and Puerto Rico had a 10-nt deletion starting at the 19th nucleotide position from the ORF termination codon (13).
The terminus of the 3´ NCR has a conserved sequence and secondary structure. The functions of the 3´ NCR of flaviviruses have not been fully determined. The 3´ NCR in flaviviruses affects RNA replication but does not affect viral translation (14,15). Introduction of a 30-nt deletion starting at the 212th position from the ORF termination codon in the 3´ NCR of DENV-4 reduced the ability of the virus to propagate in vivo and in vitro (16).
We have identified a 29-nt deletion in the 3´ NCR of DENV-1 isolated from a dengue patient returning to Japan from Yap. Isolates from 3 other patients infected in the same outbreak also had the same deletion. The DENV-1 strain with a 29-nt deletion in the 3´ NCR was responsible for the dengue epidemic in Yap in 2004. The biologic characteristics induced by this deletion should be further analyzed.
Dr Nukui is a graduate student in the Department of Infectious Diseases, University of Tokyo, Tokyo, Japan. Her research interest is vectorborne viral diseases.
Acknowledgments
We thank doctors from the various clinics and hospitals for providing us with serum samples for laboratory confirmation of dengue infection.
This work was supported by a grant for research on emerging and reemerging infectious diseases from the Ministry of Health, Labor, and Welfare, Japan.
References
- Takahashi M, Yamada K, Sato Y, Ikawa K, Matsumoto Y, Sano T, Detection of dengue virus-infected patients among passengers at the quarantine station of the New Tokyo International Airport. Jpn J Infect Dis. 2002;55:215–6.PubMedGoogle Scholar
- Chambers TJ, Hahn CS, Galler R, Rice CM. Flavivirus genome organization, expression, and replication. Annu Rev Microbiol. 1990;44:649–88. DOIPubMedGoogle Scholar
- Proutski V, Gritsun TS, Gould EA, Holmes EC. Biological consequences of deletions within the 3´-untranslated region of flaviviruses may be due to rearrangements of RNA secondary structure. Virus Res. 1999;64:107–23. DOIPubMedGoogle Scholar
- Markoff L. 5´-and- 3´-non coding regions in flavivirus RNA. Adv Virus Res. 2003;59:177–228. DOIPubMedGoogle Scholar
- Savage HM, Fritz CL, Rustein D, Yolwa A, Vorndam V, Gubler DJ. Epidemic of dengue-4 virus in Yap State, Federated States of Micronesia, and implication of Aedes hensilli as an epidemic vector. Am J Trop Med Hyg. 1998;58:519–24.PubMedGoogle Scholar
- Martin B. Dengue fever type 1 outbreak in Yap. ACTION n-20 2005; 11-2. [cited 2005 Nov 14]. Available from http://www.spc.int/phs/ENGLISH/Publications/InformACTION/IA20-contents.htm
- Barrero PR, Mistchenko AS. Complete genome sequencing of dengue virus type 1 isolated in Buenos Aires, Argentina. Virus Res. 2004;101:135–45. DOIPubMedGoogle Scholar
- Tajima S, Nukui Y, Ito M, Takasaki T, Kurane I. Nineteen nucleotides in the variable region of 3´ nontranslated region are dispensable for the replication of dengue type 1 virus in vitro. Virus Res. In press. Epub 2005 Oct 24.PubMedGoogle Scholar
- Goncalvez AP, Escalante AA, Pujol FH, Ludert JE, Tovar D, Salasa RA, Diversity and evolution of the envelope gene of dengue virus type 1. Virology. 2002;303:110–9. DOIPubMedGoogle Scholar
- Gritsun TS, Venugopal K, Zanotto PM, Mikhailov MV, Sall AA, Holmes EC, Complete sequence of two tick-borne flaviviruses isolated from Siberia and the UK: analysis and significance of the 5´ and 3´-UTRs. Virus Res. 1997;49:27–39. DOIPubMedGoogle Scholar
- Nam JH, Chae SL, Won SY, Kim EJ, Yoon KS, Kim BI, Short report: genetic heterogeneity of Japanese encephalitis virus assessed via analysis of the full-length genome sequence of a Korean isolate. Am J Trop Med Hyg. 2001;65:388–92.PubMedGoogle Scholar
- Shurtleff AC, Beasley DW, Chen JJ, Ni H, Suderman MT, Wang H, Genetic variation in the 3´ non-coding region of dengue viruses. Virology. 2001;281:75–87. DOIPubMedGoogle Scholar
- Holden KL, Harris E. Enhancement of dengue virus translation: role of the 3´ untranslated region and the terminal 3´ stem-loop domain. Virology. 2004;329:119–33. DOIPubMedGoogle Scholar
- Tilgner M, Deas TS, Shi PY. The flavivirus-conserved pentanucleotide in the 3´ stem-loop of the West Nile virus genome requires a specific sequence and structure for RNA synthesis, but not for viral translation. Virology. 2005;331:375–86. DOIPubMedGoogle Scholar
- Men R, Bray M, Clark D, Chanock RM, Lai CJ. Dengue type 4 virus mutants containing deletions in the 3´ noncoding region of the RNA genome: analysis of growth restriction in cell culture and altered viremia pattern and immunogenicity in rhesus monkeys. J Virol. 1996;70:3930–7.PubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 12, Number 2—February 2006
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Ichiro Kurane, Department of Virology 1, National Institute of Infectious Diseases, 1-23-1 Toyama, Shinjuku-ku, Tokyo 162-8640, Japan; fax: 81-3-5285-1188
Top